康复患者B细胞的高通量单细胞测序鉴定出针对SARS-CoV-2的有效中和抗体
迫切需要中和抗体来有效阻止病毒进入宿主细胞,以干预COVID-19。 使用高通量单细胞RNA测序,Cao等从60例康复期患者的B细胞中鉴定出14种有效的中和抗体。 最有效的抗体BD-368-2具有高抗体 SARS-CoV-2感染小鼠的治疗和预防功效。
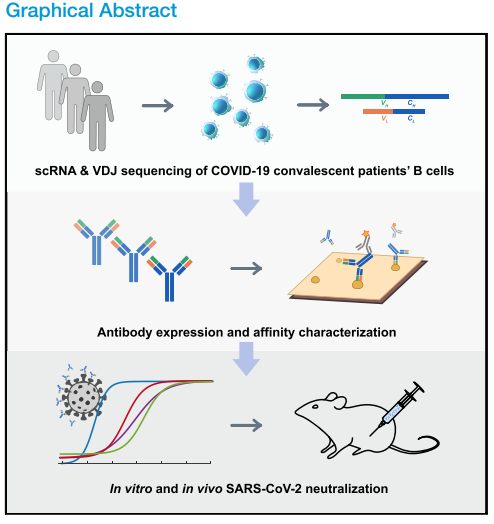
- 通过高通量scRNA / VDJ-seq鉴定了8,558个IgG1 +抗原结合克隆型
- 从60名康复中的患者中发现14种有效的SARS-CoV-2中和抗体 (怎么证明是SARS的抗体呢?)
- BD-368-2在SARS-CoV-2感染的小鼠中显示出很高的治疗和预防功效 (试验验证?)
- 中和抗体可以根据预测的CDR3H结构直接选择 (需要一些聚类吧)
COVID-19大流行迫切需要治疗和预防干预。在这里,我们报告通过高通量单细胞RNA和VDJ测序对60名康复期患者的抗原富集的B细胞进行SARS-CoV-2中和抗体的快速鉴定。从8558种抗原结合型IgG1 +克隆型中,鉴定出14种有效的中和抗体,其中最有效的一种是BD-368-2,对假型和真实(pseudotyped and authentic)SARS-CoV-2的IC50分别为1.2和15 ng / mL。 BD-368-2在SARS-CoV-2感染的hACE2转基因小鼠中也显示出强大的治疗和预防功效。另外,中和抗体与刺突-胞外域三聚体复合的3.8 A cryo-E结构显示抗体Ys表位与ACE2结合位点重叠。以前,我们证明可以根据预测的CDR3H结构与SARS-CoV中和抗体的相似性直接选择SARS-CoV-2-中和抗体。总之,我们表明可以通过高通量单B细胞测序有效地发现人中和抗体,以应对大流行性传染病。
一、前言
由一种名为严重急性呼吸系统综合症冠状病毒2(SARS-CoV-2)的新型冠状病毒引起的冠状病毒疾病2019(COVID-19),已作为一种严重的大流行病在世界范围内传播(Callaway等,2020)。 SARS-CoV-2和SARS-CoV均属于乙型冠状病毒属的谱系B(Zhou等,2020; Wu等,2020),其RNA基因组具有约82%的同一性(Chan等,2020) )。 SARS-CoV-2感染靶细胞的机制已得到充分研究,最近已报道(Hoffmann等,2020; Walls等,2020)与SARS-CoV相似,SARS-CoV-2表面的刺突(S)糖蛋白介导病毒的膜融合和受体识别(Wrapp等人,2020)。 N端区域的S1亚基负责病毒附着,并包含受体结合域(RBD),该域直接与宿主细胞上的ACE2受体结合。 目前,尚无针对病毒靶标相互作用的经过验证的疗法可用于COVID-19。
康复期患者的血浆含有适应性免疫反应产生的中和抗体,当用作治疗手段时,已导致轻度和重度COVID-19患者的明显临床改善(Chen等人,2020年; Shen等人, 2020;曹,2020)。但是,由于不能大规模生产血浆,因此治疗用途受到限制。另一方面,由于其可扩展性和治疗效果,从恢复期患者的记忆B细胞中分离出的中和单克隆抗体(mAb)可作为SARS-CoV-2的有前途的干预手段。针对病毒表面蛋白的人源单克隆抗体日益显示出其对诸如HIV,埃博拉病毒和中东呼吸综合征(ERS)等传染病的治疗和预防功效(Corti等人,2016; Wang等人,2018; Scheid等人等(2009)。他们在患者中的安全性和效力已在多项临床试验中得到证实(Xu等人,2019; Caskey等人,2017)。尽管具有优势,但从人类记忆B细胞中筛选有效的中和mAb往往是一个缓慢而费力的过程,在应对全球范围的健康突发事件时,这并不理想。迫切需要一种快速有效的方法来筛选SARS-CoV-2-中和单克隆抗体。
由于VDJ重组和体细胞超突变,B细胞表现出多样化的B细胞库,需要一次分析一个B细胞(Bassing等,2002)。记忆B细胞的单细胞克隆扩增等技术通常用于从异种B细胞群体中获得配对的免疫球蛋白重轻链RNA序列以产生mAb(El Debs等人,2012; Niu等人, 2019)。已证明从恢复期患者的爱泼斯坦-巴尔病毒(EBV,Epstein-Barr virus)永生化记忆B细胞的克隆扩增成功分离出针对病毒感染(例如HIV,登革热和ERS)的中和单克隆抗体(Burton等,2009; Corti等,2015 ; Scheid等人,2009; Setthapramote等人,2012)。然而,由于费时的孵育和筛选步骤,该技术至少需要几个月才能完成成功的筛选。
另一方面,单细胞RT-PCR结合荧光激活细胞分选(FACS)或光流体平台(例如Beacon(Berkley Light)),可以通过在单个抗原结合记忆体上执行嵌套式PCR在几天内获得抗体序列单细胞分选后的B细胞(Tiller等人,2008; Wardemann等人,2003; Wrammert等人,2008; Liao等人,2009)。该方法已导致在包括HIV和ERS在内的各种传染病中有效分离中和mAb(Scheid等,2009; Wang等,2018)。尽管如此,最近在使用BX筛选效率方面,使用10X Chromium对B细胞受体库进行高通量单细胞RNA和VDJ测序的最新进展已经超过了单细胞RT-PCR(Goldstein等人,2019; Horns等人) 。,2020)。基于微流体的技术可以一次从成千上万的单个B细胞中获得自动配对的重链和轻链序列,并已成功用于分离针对HIV的人中和单克隆抗体(Setliff等,2019)
在这里,我们报告通过恢复期COVID-19患者的抗原结合B细胞的高通量单细胞RNA和VDJ测序,实现的SARSCoV-2中和抗体的快速有效鉴定。从60名康复中的患者中鉴定出超过8,500种表达免疫球蛋白G1(IgG1)抗体的抗原结合B细胞克隆型。我们总共鉴定出14种有效的中和mAb,其中最有效的mAb BD-368-2对假型和真实SARS-CoV-2的IC50为1.2和15 ng / mL。此外,体内实验证实,使用hACE2转基因小鼠模型,BD-368-2可提供强大的治疗功效和针对SARSCoV-2的预防性保护(Bao等,2020; Yang等,2007; cCray等。 ,2007)。
此外,我们解决了与SARS-CoV-2穗胞外域三聚体(spike ectodomain trimer )复合的一个中和单克隆抗体BD-23的低温电子显微镜(cryo-E)结构,并显示其表位与RBD / ACE2结合基序重叠。基于SARS-CoV-2和SARS-CoV之间的高度保守性,我们证明,可以利用CDR3H结构的相似性,直接从大型抗原结合克隆型文库中直接选择针对SARS-CoV-2的有效中和单克隆抗体。 SARS-CoV中和抗体 m396 的制备(Prabakaran等,2006; Zhu等,2007)。总体而言,我们表明,高通量单细胞测序可导致鉴定具有强大的治疗和预防功效的高效中和单克隆抗体,这可极大地协助干预流行和新兴传染病,例如COVID-19。
二、结果
2.1 康复患者单B细胞的高通量测序 High-Throughput Sequencing of Single B Cells from Convalescent Patients
与传统方法不同,从高通量scVDJ测序(scVDJ-seq)获得的大规模数据使我们能够在体外抗体表达之前检查B细胞克隆型富集情况(Goldstein等,2019; Croote等,2018)。 对于重链和轻链均具有相同CDR3区域的B细胞被分组为相同的克隆型(clonotypes),并根据观察到的克隆型细胞数计算其富集度。 由于抗原激活的B细胞会经历克隆选择并从已有的幼稚和记忆B细胞中扩增(Murugan等人,2018; Seifert和Küppers,2016),因此我们假设富集的B细胞克隆型更有可能产生高 -亲和性SARS-CoV-2结合和中和抗体。
为了利用这一假设,我们首先收集了北京尤安医院的12名COVID-19恢复期患者的外周血单核细胞(PBMC)并分离了B细胞(表S1)。我们使用10X Chromium 5’mRNA和VDJ测序对新鲜分离的B细胞或CD27 +记忆B细胞亚群进行了小条件RNA(scRNA)和scVDJ测序(图1A和S1A)。 scVDJ数据确实显示了丰富的IgG1克隆型(图S1F和S1H),而CD27 +记忆B细胞的选择大大改善了测序的记忆B细胞的数量(图S1D和S1E)以及发现的IgG1克隆型(图S1G和S1I)。 。但是,从包含丰富的IgG1 +克隆型的记忆B细胞中选择的130种体外表达的单克隆抗体(表S2)中,两种命名为BD-23的RBD结合单克隆抗体中只有一种显示出弱的病毒中和能力,无法抵抗假型和真实SARS-CoV-2(图S1B和S1C)。在RBD结合mAb和中和mAb识别方面效率低下,因此需要基于抗原亲和力的选择,这可能会极大地增强掺入刺突/ RBD的B细胞。
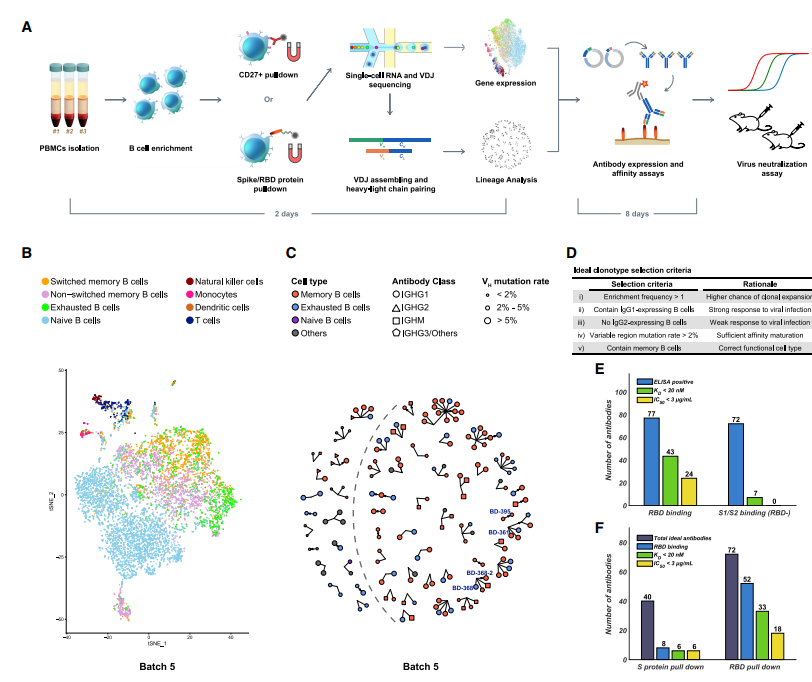

2.2 通过抗原结合单B细胞的高通量测序高效鉴定中和单克隆抗体 Efficient Identification of Neutralizing mAbs by High-Throughput Sequencing of Antigen-Binding Single B Cells
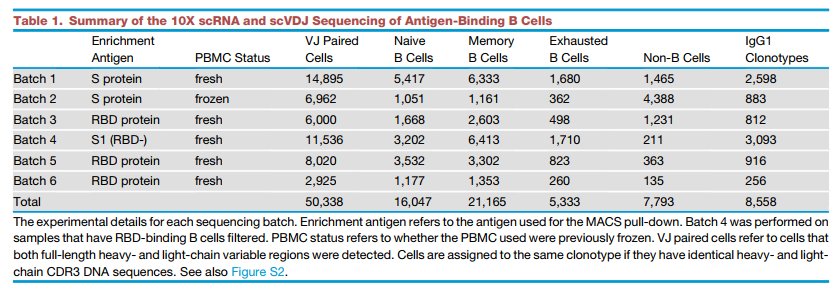
为了丰富结合RBD的B细胞,我们利用生物素化的RBD和S蛋白通过磁珠分离(图1A)来选择结合抗原的B细胞。由于选择会导致B细胞数量的大幅减少,因此我们组合了来自不同患者的PBMC,以提供足够的10倍负荷量。我们总共分析了6个不同批次的60例康复期患者的抗原结合B细胞(表S1)。由于目标是产生中和单克隆抗体,因此我们滤除了所有无法产生跨越宽轻链对的V-J的B细胞。总共检测到8,558种不同的IgG1呈递抗原结合克隆型(表1)。此外,scRNA-seq数据使我们能够基于mRNA表达进行细胞分型,以鉴定记忆B细胞(图1B和S2)。我们观察到幼稚B细胞和记忆B细胞之间以及非B细胞与记忆B细胞之间有足够的分离。但是,在某些批次中可能未观察到开关/非开关记忆B细胞与耗尽记忆B细胞之间的清晰分离,这主要是由于相对较低的测序深度以及scRNA测序的每个细胞检测到的基因数量较少(表S3)。
为了提高中和单克隆抗体鉴定的效率,我们制定了一套标准,以筛选出产生中和单克隆抗体可能性低的克隆型(图1D)。
- 首先,仅选择含有表达IgG1的B细胞的富集克隆型(图1C和S3A),因为表达IgG1的B细胞对病毒刺激反应强烈(Vidarsson等,2014)。
- 其次,表达IgG2的B细胞通常对病毒病原体反应较弱,因此不包括呈IgG2的克隆型作为理想候选基因(Irani等人,2015)。
- 接下来,不含任何B细胞的体细胞超突变率(SHM)高于2%的克隆型表明亲和力成熟不足,因此被排除(Croote等人,2018; Methot和Di Noia,2017)。
- 最后,由于精疲力竭的B细胞和幼稚B细胞对抗原刺激的反应较弱(Moir和Fauci,2014),因此仅包含精疲力竭的(exhausted )记忆B细胞或幼稚B细胞的克隆型不被视为理想候选者(图1B和S2)。
一起,从富集的克隆型中总共选择了169种理想候选物,并通过转染在HEK293细胞中表达(表S2)。同时,在体外还表达了47个非理想的候选人,以验证我们选择标准的有效性(表S2)。
Figure S3
通过ELISA和表面等离子体共振(SPR)测试了纯化的mAb的SARS-CoV-2 RBD /spike 反应性,并且鉴定了149种S结合mAb,其中超过一半的mAb与RBD结合。我们进一步使用SARS-CoV-2伪病毒系统筛选了所有ELISA阳性mAb的中和能力。我们发现只有结合RBD的mAb才显示出伪病毒中和作用(图1E),只有结合到RBD且其KD小于或接近ACE2 / RBD的解离常数(15.9 nM)的mAb(图S5B)才具有显着性对SARS-CoV-2假病毒的中和作用(IC50 <3μg/ mL)(图S4A和S4B)。全长S蛋白和RBD蛋白均可富集RBD结合mAb,但效率却不同(图1F)。使用RBD蛋白作为富集抗原导致RBD ELISA阳性率比使用S蛋白高3倍。最重要的是,从理想候选物中选择的单克隆抗体在鉴定强结合RBD的单克隆抗体(KD <20 nM)和SARS-CoV-2中和单克隆抗体上分别显示出46%和25%的效率(图S3D)非理想单克隆抗体候选物(图S3B和S3C)。
Figure S5
Figure S4
2.3 丰富克隆型单克隆抗体对SARS-CoV-2的高中和力 High Neutralizing Potency against SARS-CoV-2 among mAbs from Enriched Clonotypes
从富集的克隆型中分离出的所有中和mAb中,有7种显示出有效的中和能力,对SARS-CoV-2假病毒的IC50低于0.05μg/ mL(图2A和2C)。从与BD-368相同的克隆型中选择的最有效的mAb BD-368-2(图1C)显示出IC50为1.2 ng / mL。 SPR显示,所有七个mAb均以nM或亚nM KD与RBD牢固结合(图S5A),并且可以竞争性抑制ACE2与RBD结合(图S5C和S5D)。为了评估其对真病毒的中和潜力,我们使用了从COVID-19患者中分离出的真SARS-CoV-2进行了噬斑减少中和测试(PRNT)。在所有测试的单克隆抗体中,BD-368-2表现出对真病毒的最高效力,IC50为15 ng / mL(图2B和2C)。为了进一步验证PRNT显示的真实病毒中和作用,我们使用BD-218对真实SARS-CoV-2进行了细胞病变效应(CPE)抑制测定。 BD-218在所有三个重复实验中均表现出完全的病毒抑制作用,浓度为1.2μg/ mL(图S6A和S6B),与PRNT结果一致。总之,我们的数据表明,通过抗原结合B细胞的高通量单细胞测序,可以从恢复期患者中鉴定出高效的中和mAb。
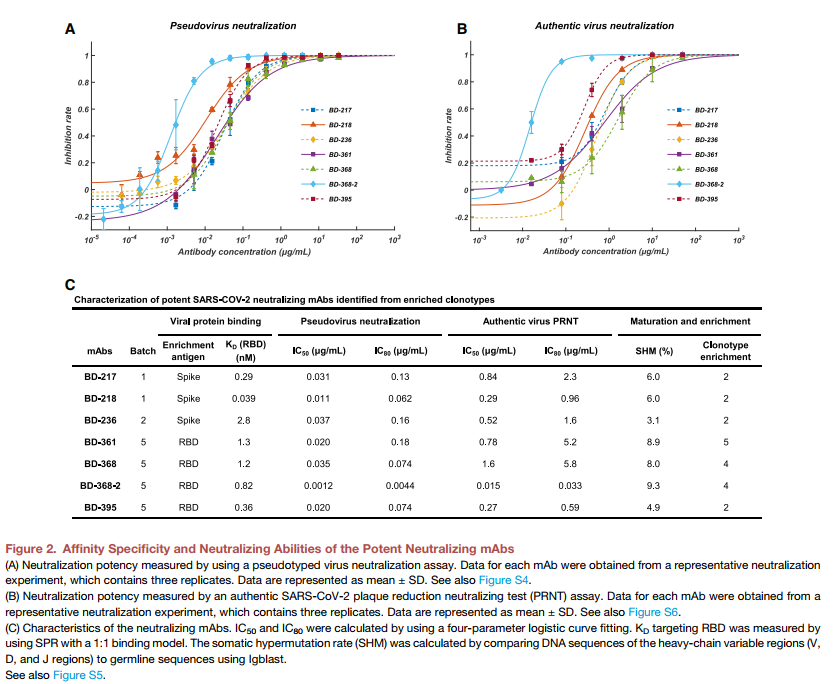
Figure S6.
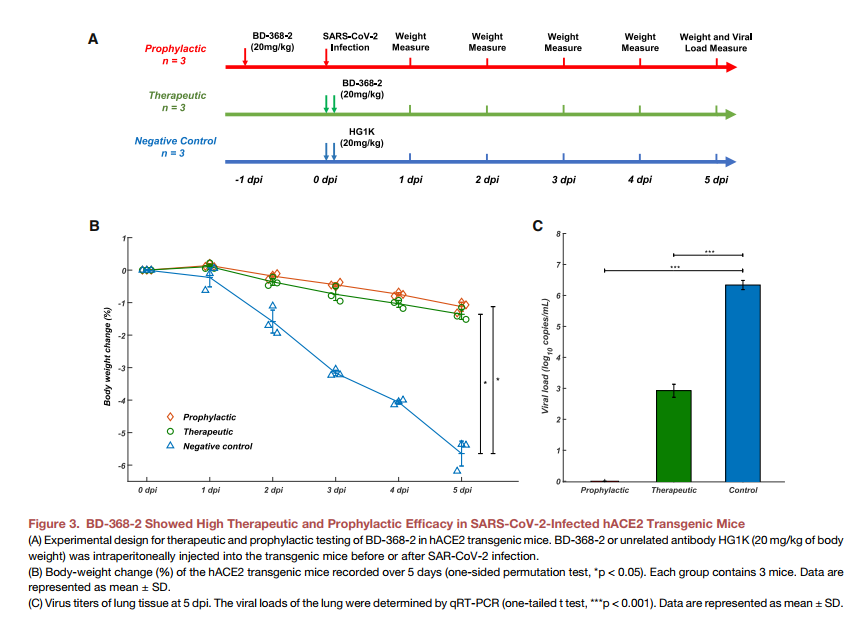
2.4 BD-368-2在hACE2小鼠中显示出很高的治疗和预防功效
为了评估鉴定出的中和单克隆抗体是否可以在体内作为针对SARS-COV-2的治疗性干预措施和预防性保护措施,我们测试了BD-368-2对感染SARS-CoV-2的hACE2转基因小鼠的中和功效(Bao等。,2020)。之所以选择BD-368-2,是因为它对假型和真实SARS-CoV-2都表现出最高的效力。这项研究总共使用了9只hACE2转基因小鼠,并将它们平均分为三组(n = 3)。通过在病毒感染前24小时向hACE2转基因小鼠腹膜内注射20 mg / kg(小鼠体重)BD-368-2来检查预防效果(图3A)。对于治疗组,感染后2小时注射20 mg / kg BD-368-2。作为阴性对照,在感染后2小时注射20 mg / kg HG1K,一种针对H7N9病毒的IgG1抗体。我们在感染5天后每天记录每只小鼠的体重(图3A),发现治疗组和预防组均保持了体重,而阴性对照组的体重明显减轻(图3B)。这表明在感染之前和之后均施用BD-368-2可以极大地改善SARS-CoV-2感染小鼠的生理状况。此外,我们通过qRT-PCR分析了5 dpi时肺部的病毒载量(Yang等,2007),发现在病毒攻击之前注射BD-368-2可以完全抑制SARS-CoV-2(图3C) )。此外,在感染后2小时应用BD-368-2可能导致小鼠肺部病毒复制减少3-4 log,这表明SARS-CoV-2复制的有效减少(图3C)。一起,BD-368-2在体内表现出高的治疗和预防功效。
2.5 绑定到Trimeric Spike蛋白的中和单克隆抗体的低温-EM结构。
在分析了丰富的克隆型后,可以有效地鉴定出针对SARS-CoV-2的中和单克隆抗体。但是,由于每位患者的B细胞采样较浅,大多数克隆型均未显示任何富集。尽管如此,未富集的抗原结合克隆型仍保留为有价值的文库,用于将来鉴定中和mAb。需要一种针对未富集克隆型的新抗体选择标准,因为任意选择可能会导致高百分比的弱抗原结合单克隆抗体,并且在抗原下拉过程中也容易发生非特异性结合。以前,人们使用抗体结构预测来注释Ig-seq数据,以更好地选择mAb(DeKosky等,2016)。通过选择与参考中和mAb共享相似CDR3H结构的mAb,结构注释方法已被证明可成功改善Ig-seq的性能以及体外抗体成熟(Kovaltsuk等人,2017; Krawczyk等人。 (2014年,Sela-Culang等人,2014年)。受这些结果的启发,我们假设基于CDR3H的结构预测也可以帮助注释来自恢复期患者的scVDJ-seq数据,这可能有助于提高针对SARS-CoV-2的中和单克隆抗体的识别效率。
然而,主要困难在于参考抗体结构的选择。迄今为止,蛋白质数据库(PDB)中未记录任何SARS-CoV-2中和mAb结构,可作为参考。为了解决这个问题,我们解决了BD23-Fab与Sectodomain三聚体复杂的低温EM结构,总分辨率为3.8Å(图4A和S7;表S4)。这里的Sectodomain采用先前报道的不对称构象(Wrapp等,2020),其中一个Protomer(mol A)中的RBD处于“ up”位置,而其他两个RBD(mol B和C)则采用“下”的位置。在此3D重建中,每个S三聚体均观察到单个BD23-Fab,并且它与原型B中的“向下” RBD结合。只有BD-23的重链可变域与RBD结合(图4B)。有趣的是,还可以通过在启动子C的Asn165上的N-连接聚糖来促进结合。尽管当前的分辨率不支持对BD23-Fab和RBD之间的分子相互作用的进一步分析,但很明显BD23的表位与公认的基序重叠通过ACE2。实际上,与RBD-ACE2复杂结构的比较表明BD23-Fab与ACE2竞争与RBD的结合(图4C)。该观察结果表明,BD-23的SARS-CoV-2中和能力来自ACE2-RBD结合的破坏,这与先前鉴定出的针对SARS-CoV的中和单克隆抗体高度相似(Prabakaran等,2006,Yuan等)等(2020年)。实际上,一些研究表明,中和SARS-CoV的单克隆抗体可以交叉中和SARS-CoV-2(Wang等,2020; Tian等,2020; Lv等,2020),这进一步突显了相似之处。在SARS-CoV和SARS-CoV-2的RBD之间。总的来说,SARS-CoV和SARS-COV-2之间的结构观察和高度保守性表明,使用SARS-CoV中和单克隆抗体的晶体结构作为参考来筛选SARS-CoV-2抗原也是可行的。共享相似的CDR3H结构的结合型克隆型,以鉴定中和mAb。
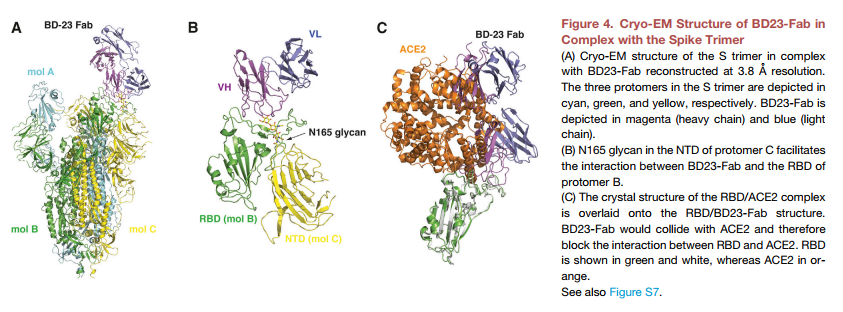
Figure s7
2.6 基于CDR3H的结构相似性选择有效的中和单克隆抗体 Selecting Potent Neutralizing mAbs Based on the Structural Similarity of CDR3H
我们将使用FREAD基于CDR3H的每个IgG1克隆型的预测结构,与PDB数据库中可用的SARS-CoV中和单克隆抗体的晶体结构以及BD-23的结构进行了比较。 不幸的是,没有IgG1的克隆型表现出与BD-23高度的CDR3H结构相似性。 但是,共有12种IgG1克隆型与PDB ID 2dd8和2ghw具有高度的结构相似性(表S2;图5C),分别对应于先前分离出的SARS-CoV中和单克隆抗体m396和80R(Prabakaran等,2006, Hwang等,2006)。 鉴定出的mAb的CDR3H序列与m396(2dd8)具有高度同源性(图5A),后者通过破坏ACE2 / RBD结合来中和SARS-CoV(图5D)。
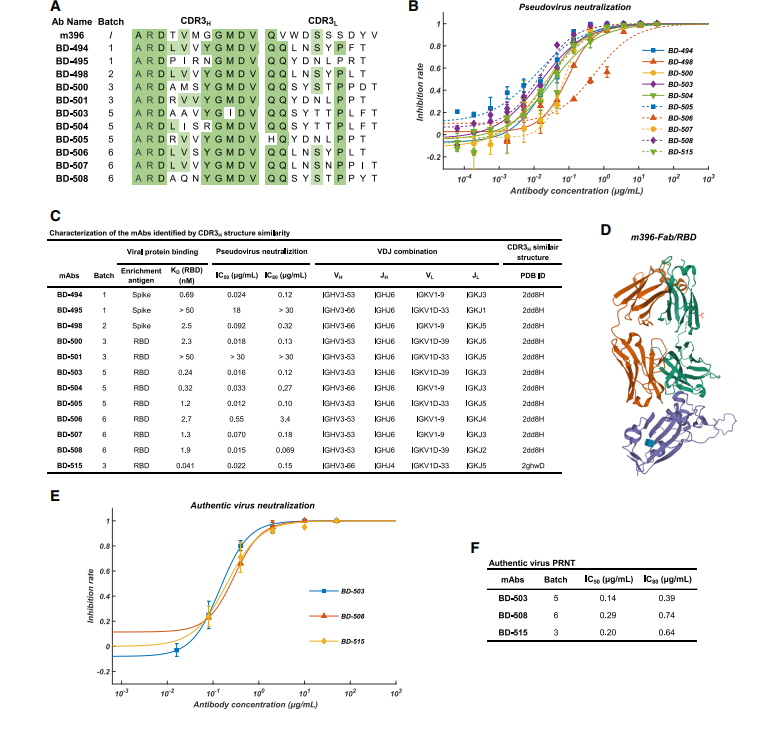

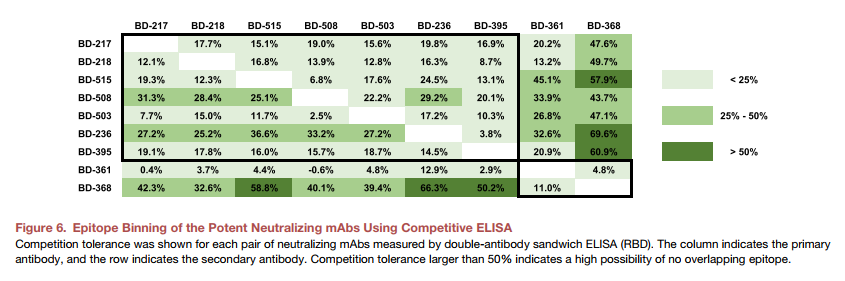
出人意料的是,我们发现12个单克隆抗体中有7个显示出强大的RBD结合亲和力和对假型SARS-CoV-2的有效中和能力(图5B和5C)。 PRNT中还针对真实的SARS-CoV-2验证了三种代表性mAb BD-503,BD-508和BD-515的中和能力,并显示出高效力(图5E和5F)。此外,这些mAb主要由重链的VH3-66 / JH6或VH3-53 / JH6基因片段编码,而轻链则由VLK1-9或VLK1D-33 / 39编码(图5C)。由于氨基酸序列和VDJ组合的相似性,我们质疑这些mAb是否来自同一谱系。经过RNA突变分析后,我们发现所有mAb都不来自同一谱系,也不来自同一位患者。总之,这些观察结果表明,与特定SARS-CoV-2表位结合的刻板B细胞受体可能存在于不同的个体中,类似于在HIV,流感和丙型肝炎中所观察到的(Gorny等,2009,Ekiert等人,2009,Marasca等人,2001)。确实,使用双抗体夹心ELISA进行的表位分箱实验表明BD-503,BD-508和BD-515可能共享重叠的表位(图6)。但是,需要更多的结构分析以获得确切的表位,以确认它们是否在很大程度上保守。
三、讨论区
为了提高发现新的中和mAb的效率,我们在进行体外表达之前应用了抗体呈递B细胞的系统分析。从scRNA和scVDJ测序获得的大规模单细胞分辨率数据提供了进行谱系分析的机会,而使用杂交瘤和基于单细胞RT-PCR的技术则无法轻松实现谱系分析。计算每种克隆型富集频率的能力不仅可以优先选择天然克隆扩增的B细胞谱系,而且还可以降低抗原富集步骤中非特异性抗原结合的假阳性率(图1A)。在单细胞水平上同时测量mRNA表达还可以实现单个B细胞的细胞分型,这对于去除疲惫和幼稚(exhausted and naive)的B细胞亚群至关重要。但是,我们目前的浅层scRNA测序几乎无法使疲惫的记忆B细胞与未疲惫的记忆B细胞准确分离。因此,需要对scRNA文库进行更深的测序,以进一步评估去除耗尽的记忆B细胞以改善中和mAb鉴定的有效性。
除克隆富集的B细胞克隆型外,绝大多数测序的B细胞未在本研究中使用,但仍作为潜在的中和mAb的宝贵文库。然而,由于缺乏富集,从大量未富集的B细胞中选择mAb仍然是一个问题。因此,我们利用了基于生物信息学的选择方法,例如CDR3H结构预测。与SARS-CoV中和抗体m396共享高度相似的CDR3H结构的mAb对SARS-CoV-2的高中和效能显示出令人惊讶的高百分比。 VDJ基因片段和这些mAb可能重叠的表位的高度保守组合表明存在针对SARS-CoV-2的原型B细胞受体。根据不同个体的VH3-66 / JH6或VH3-53 / JH6编码的重链选择的mAb的初步研究确实显示,高比例的SARS-CoV-2中和单克隆抗体具有很高的效价(数据未显示),并支持存在对SARS-CoV-2的定型B细胞受体。
尽管已鉴定出72个S1 / S2(非RBD)结合mAb,但它们均未显示出伪病毒中和能力。这是一个有趣的结果,因为在MERS-CoV中观察到了非RBD结合中和mAb(Xu等人,2019)。从针对抗原不同区域的抗原结合B细胞克隆型的剩余库中筛选更多抗体的中和能力,以进一步证实观察结果。此外,更多的伪病毒株可用于检查病毒突变后中和逃逸的潜在风险,尤其是在RBD区(Wang等,2018)。病毒突变的一种可能解决方案是将两种针对两个不同表位的有效mAb结合在一起。表位合并结果(图6)表明,与BD-368-2共享相同表位的BD-368,在大多数有效的中和单克隆抗体上均表现出不重叠的表位。与任何单个mAb相比,将BD-368-2与另一种中和mAb结合的抗体混合物可以大大提高SARS-CoV-2中和能力,并且可以很大程度上防止突变逃逸,因为不太可能在两个表位上都出现突变。
作为全球大流行的持续传播,COVID-19要求对SARS-CoV-2感染采取有效的干预措施。我们鉴定出的有效中和抗体可能提供有效的治疗和预防解决方案,这在hACE2转基因小鼠的研究结果中得到了证明。正在使用BD-368-2进行临床试验。高通量单细胞测序方法在本研究中的应用还可以扩展到COVID-19以外的其他传染病,并为在未来大流行期间快速中和mAb的发现提供技术储备。
四、方法
试验略
4.1 B细胞scRNA-seq数据分析
- 使用10X Genomics CellRanger(3.1.0)管道处理原始的fastq文件。 将5’基因表达谱产生的读数与GRCh38基因组进行比对,并使用带有默认参数的“ cellranger count”生成特征条形码矩阵。 为了滤除低质量的基因和细胞,保留了超过10个细胞中表达的基因,并根据基因数量和线粒体基因百分比对细胞进行了过滤,以去除可能的双峰。
- 使用SingleR(Aran等人,2019)针对人类免疫参考数据集(Monaco等人,2019)鉴定细胞类型。
- 使用Seurat中的T分布随机邻居嵌入(t-SNE)可视化细胞簇(Satija等人,2015)。
4.2 B细胞scVDJ数据分析 B cell scVDJ data analysis
使用10X Genomics CellRanger(3.1.0)管道处理原始fastq文件。 质量得分低于30的碱基的3’端使用cutadapt修剪(Martin,2011)。 重叠群的组装,注释和克隆型分析是使用“ cellranger vdj”和Cell Ranger V(D)J兼容的参考文献进行的:
(refdata-cellranger-vdj-GRCh38-alts-ensembl-3.1.0)。
4.3 BCR曲目的结构注释 Structural annotation of BCR repertoires
重链和轻链中三个CDR区的结构使用SAAB +管道(Kovaltsuk等人,2020)和默认参数进行注释。 SAAB +利用SCALOP(Wong等人,2019)来识别CDR1和CDR2序列的典型类别。 CDR3序列的结构注释是通过整合在SAAB +流水线中的FREAD(Choi和Deane,2010)完成的。
4.4 体细胞超突变率测定
V(D)J测序读数的映射是由IgBlast-1.15.0(Ye等人,2013)针对从国际ImMunoGeneTics信息系统(IMGT)下载的种系参考文献(202007-1版本)(Giudicelli等人) (2006年)。 高变异率的计算公式为,其中缺口的计算公式为插入或缺失区域中碱基对的数量。
(mismatches +gaps) / length of the query sequence
4.5 理想的抗体选择
提取所有属于富含IgG1克隆型的B细胞,并记录其相应的细胞类型,体细胞超突变率(SHM)和Ig类。 理想的抗体选择过程严格遵循图1D中列出的标准。 每个克隆型的血统都由DNA突变模式以及Ig类决定。 谱系关系不会干扰理想的抗体选择,只会在从特定克隆型中选择mAb序列时有所帮助。 当相同克隆型的B细胞不共享相同的VDJ序列时,通常选择谱系中最成熟的B细胞的DNA序列进行体外表达。 世系(lineage)图由igraph绘制(Csardi和Nepusz,2006年)。 图中单元格的位置没有物理意义,可以任意调整以将所有理想克隆型分组到一侧。
问题
问题1:
我们使用10X Chromium 5’mRNA和VDJ测序对新鲜分离的B细胞或CD27 +记忆B细胞亚群进行了小条件RNA(scRNA)和scVDJ测序(图1A和S1A)
scRNA 和 scVDJ测序的目的是什么? 10X Chromium的有点是啥?
参考资料
- Cao et al., 2020, Cell 182, 73–84 July 9, 2020 ª 2020 Published by Elsevier Inc. https://doi.org/10.1016/j.cell.2020.05.025
