【8.5.3】随机肽文库显示在腺相关病毒上以选择靶向基因治疗载体
表征细胞表面的分子多样性对于靶向基因治疗至关重要。细胞类型特异性结合配体可用于靶向基因治疗载体。然而,不能获得其中可以在感兴趣的细胞上选择最佳真核载体的靶向系统。在这里,我们介绍并验证随机腺相关病毒(AAV)肽库,其中每个病毒颗粒在衣壳表面显示随机肽。该文库在三步系统中生成,该系统确保通过包装的DNA编码展示的肽。作为概念验证,我们筛选了人冠状动脉内皮细胞上的AAV文库。我们观察到特定肽基序的选择。所选择的肽增强冠状动脉内皮细胞的转导,但不控制非内皮细胞的转导。这种载体靶向策略优于其他组合方法,例如噬菌体展示,因为选择发生在衣壳的背景下,并且可能在生物技术和医学中具有广泛的应用。
一、前言
与当前基因治疗载体相关的许多安全性和功效问题,可以通过消除其内源性不希望的向性并将它们重新靶向特定组织来克服。 为了将病毒载体靶向细胞类型特异性受体,配体必须通过化学(双特异性缀合物)或重组(基因修饰衣壳)方法与载体的帽表面连接。 随机噬菌体展示肽文库可用于鉴定与某些细胞类型结合的配体。 此类配体已用于实验模型中的治疗靶向
通过含有靶向肽的双特异性缀合物重定向病毒载体对于全身治疗具有显着缺点。 这些包括体内衔接子 - 载体复合物的不稳定性和衔接子分子的免疫原性。 因此,通过将配体直接掺入其病毒衣壳中来重新定位载体是优选的。 通过噬菌体展示选择的肽掺入病毒衣壳中的这种掺入可能是成功的,但也具有局限性。 肽构象可以在病毒蛋白质环境中改变,并且配体 - 受体结合特异性和亲和力可以降低或丧失。 此外,如果相应的受体不内化(internalize)配体或以不支持基因转导的方式将其内化,则通过噬菌体展示分离的肽可能不能有效地用于基因治疗载体靶向目的。
为了克服这些限制,我们设计了一种直接在基因治疗载体上展示的肽筛选系统,即重组腺相关病毒2型(AAV-2)。 AAV载体是有吸引力的,因为它们是非致病性的,不依赖于促进转导细胞并在体内实现长期转基因表达。硫酸乙酰肝素蛋白多糖(Heparan sulfate proteoglycan)作为AAV-2的主要附着受体。已提出人成纤维细胞生长因子受体-1和αVβ5整联蛋白作为二级受体。单链AAV基因组的两侧是反向末端重复序列(ITR,inverted terminal repeats),其中含有复制和包装所需的顺式元件。 cap基因编码三种衣壳蛋白VP1-3。已经绘制了抗原区域以及AAV-2衣壳的肝素结合位点。此外,已经确定了适合掺入靶向肽的AAV衣壳内的几个位点。将合适的肽插入VP1-3的环IV中将AAV-2载体重新靶向对野生型AAV-2不敏感的细胞。最近已经解决了AAV-2衣壳的结构,为合理设计新的配体插入位点奠定了基础。
在这些研究的基础上,我们设计了一个在AAV-2表面显示的随机肽库。 将文库肽定位成使得它们在对于病毒附着于靶细胞至关重要的位点暴露于载体衣壳的表面,从而减少载体的非特异性内源性向性。 这些文库用于选择感染原代人冠状动脉内皮细胞的AAV。 选择的衣壳共享共同的肽基序和介导的冠状动脉内皮细胞类型特异性转导优于携带野生型衣壳的对照载体。 我们的研究结果可能允许选择特定的载体衣壳,以针对任何给定的感兴趣细胞类型进行基因治疗。
二、结果
2.1 AAV-2肽文库质粒的产生和评估
我们设计了一个基于pMT187-0-3质粒的AAV-2文库,在VP衣壳蛋白结构域内显示随机的七个氨基酸肽,这是AAV-2与其天然受体硫酸乙酰肝素结合所必需的(图1a)。随机肽的两侧是两个固定的氨基酸,甘氨酸和丙氨酸(图1b)。产生的文库质粒的多样性是每个文库1.1×108个克隆。 100个克隆的DNA测序证实在每个克隆中编码不同的随机肽(数据未显示)。将具有插入LHLNVAL或AALAGSL的这些克隆中的两个,以及无插入骨架质粒用于病毒生产。设计文库骨架使得需要寡核苷酸插入以将阅读框移回原始类型,如野生型cap基因。因此,无插入文库骨架质粒pMT187-0-3不产生AAV颗粒, 而克隆LHLNVAL或AALAGSL使用pSub201野生型质粒从AAV-2产生的颗粒的基因组和衣壳滴度在30%至70%之间。 与克隆LHLNVAL或AALAGSL产生的结合肝素的AAV,相对于野生型AAV-2减少70-90%(数据未显示)。 这些数据表明,cap基因内的突变和插入允许完整的含有基因组的AAV的有效组装,但是减少它们与肝素的结合。
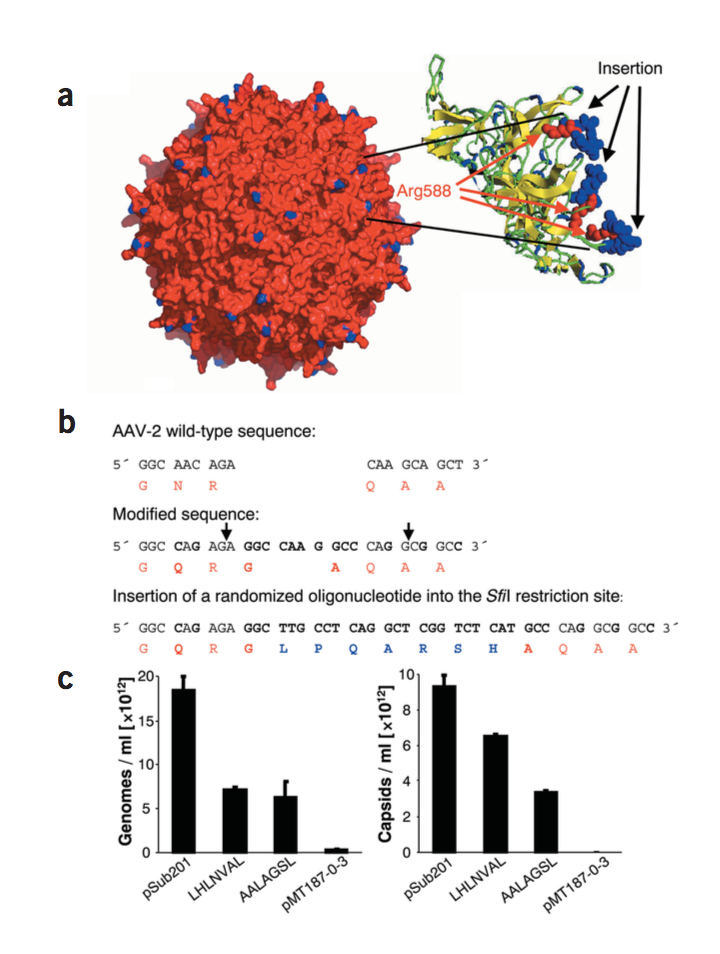
图1 随机肽AAV展示文库构建体的设计和特征。 (a)AAV-2衣壳的拓扑结构。蓝色,7个额外随机氨基酸残基的位置接近AAV-2衣壳表面上三重尖峰的顶部。插入物显示穿过较高分辨率的刺突区域的横截面,其中Arg588显示为红色,而相邻的七个氨基酸源自显示为蓝色的文库插入。 (b)在核苷酸位置3967的诱变之前和之后,AAV-2基因组内的cap基因的一部分,以获得文库骨架质粒(pMT187-0-3)。新质粒含有两个SfiI限制性位点(用箭头标记)。这两个位点由’填充'(stuffer)寡核苷酸分开。如果该填充物未被合适的寡核苷酸插入物替换,则读取框移位一个核苷酸,从而防止在没有插入物的情况下产生AAV。核苷酸序列是黑色字母。与野生型序列相比的变化以粗体突出显示。各氨基酸残基序列(单字母代码)为红色字母。蓝色字母表示插入编码肽LPQARSH的随机寡核苷酸的另外7个氨基酸残基。允许野生型衣壳氨基酸从天冬酰胺到谷氨酰胺的变化,因为预计它会导致衣壳内的空间变化最小。克隆到该衣壳区域的插入物由两个新的小氨基酸甘氨酸和丙氨酸构成,因为编码这些氨基酸的核苷酸序列是限制性位点所必需的,并且甘氨酸和丙氨酸充当文库肽插入物与周围衣壳蛋白之间的间隔区。 。 (c)对VP衣壳蛋白内七个氨基酸肽插入物产生的AAV颗粒数量的影响。如所述产生具有野生型或突变衣壳的病毒(pSub201质粒用于产生野生型AAV,空pMT187-0-3质粒用作对照)。通过斑点印迹分析和随后的磷成像读数定量的纯化的基因组AAV表示为基因组/ ml(左);通过A20-衣壳ELISA定量的AAV衣壳表示为衣壳/ ml(右)。对照值处于背景水平。值是平均值+ s.d
2.2 生成随机AAV-2展示肽库
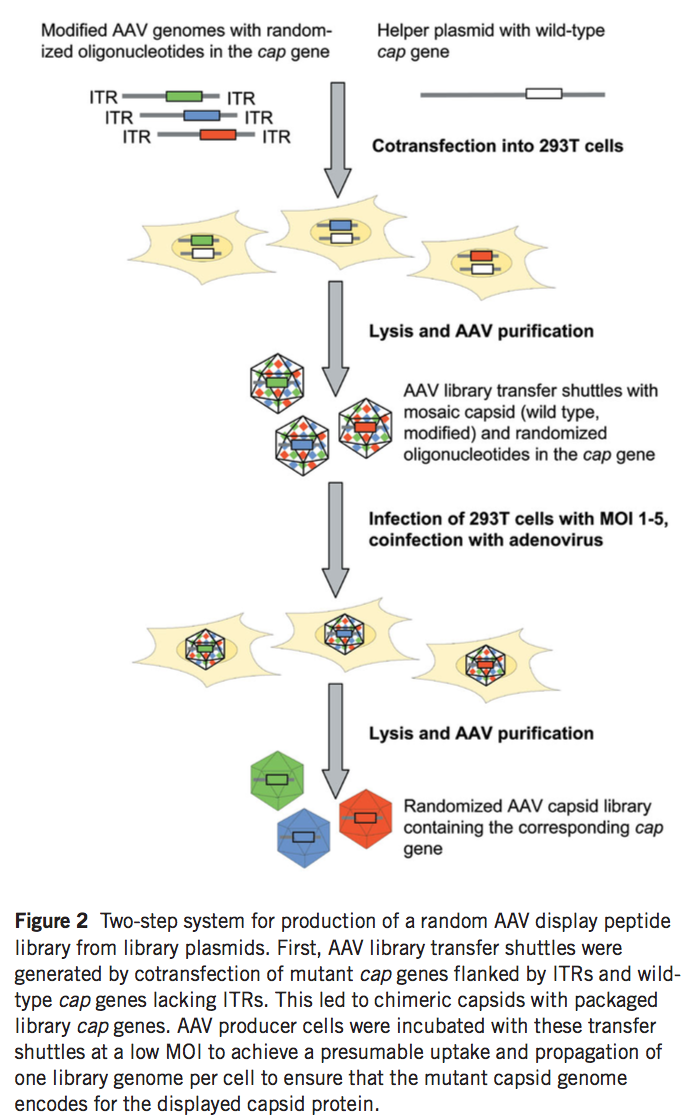
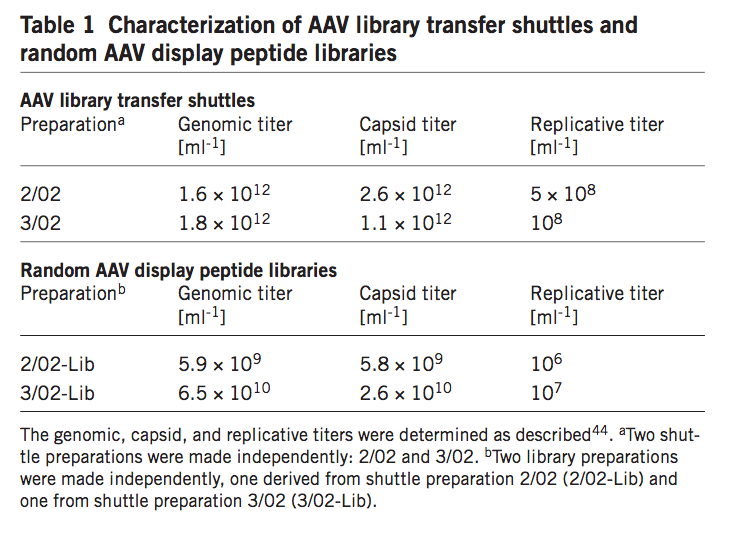
在建立质粒文库后,在另外两个步骤中进行AAV颗粒文库(AAV particle library)的产生(图2)。我们推断,通过使用常规DNA转染程序将质粒文库直接转移到AAV生产细胞中,可能导致每个生产细胞摄取超过一个文库质粒,随后产生显示多于一种类型肽插入物的嵌合衣壳。 为了避免这个问题,我们首先将含有突变型cap基因的文库打包成部分由野生型VP蛋白组成的衣壳, 通过共转染缺乏衣壳化所需的ITR的野生型rep-cap质粒(pXX2)以及AAV质粒文库。 得到的AAV颗粒,称为’AAV文库转移穿梭机',是在高基因组和复制滴度下产生的(表1)。穿梭颗粒以约60%野生型AAV-2的水平结合肝素(在线补充表1)。接下来,用AAV文库转移穿梭机以每个细胞1个和5个复制单位的感染复数(MOI)感染293T细胞,从而估计每个细胞约1-5个文库基因组的摄取和繁殖。用野生型5型腺病毒(Ad5)对细胞进行超感染,导致随机AAV-2展示肽文库的有效产生(表1)。对来自该AAV展示肽文库的代表性数目的随机选择的克隆进行测序,以显示病毒文库的多样性与产生它的质粒文库中的多样性相当(补充表2和图1在线)。一种文库制剂中的肝素结合是野生型AAV-2的36%(在线补充表1)。这些结果表明AAV文库可以在含有随机寡核苷酸插入物的AAV编码质粒的两步系统中有效产生。
参考资料
-
- Random peptide libraries displayed on adeno- associated virus to select for targeted gene therapy vectors
